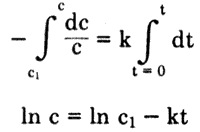(dL/g)
(dL/g)Food Technology. Vol. 38, Num. 5. May 1984. pp. 67-96.
Symposium: Gelation in Food Protein Systems
Authors Rizvi and Baker are, respectively, Associate Professor, Dept. of Food Science, and Professor and Chairmen, Dept. of Poultry and Avian Sciences, Cornell University Ithaca, NY 14853. Author Gossett formerly with Cornell, is a Scientist with The Pillsbury Co. 311 Second St. S E., Minneapolis, MN 55414
Based on a paper presented during the symposium, "Gelation in Food Protein Systems," at the 43rd Annual Meeting of the Institute of Food Technologists, New Orleans. La., June 19-22, 1984.
An important function of proteins in food systems is gelation. This phenomenon involves the formation of a three-dimensional matrix mainly through inter-protein hydrogen bonding and allows the immobilization of water within the gel structure. The coagulation or gelation of proteins—in particular, the irreversible, heat-induced coagulation or gelation of egg proteins—often controls the success of certain conked food products. It is of interest to the food scientist to be able to quantitatively monitor the gelation process to better predict end-product characteristics, as well as understand the mechanism of network formation. A better understanding of the gelation process will permit manipulation of variables to obtain a gel of desired textural characteristics and functional properties.
Although the coagulation, i.e., denaturation of proteins can be brought about by heat, high pressure, salts, acids, alkalies, alcohols, or denaturing agents such as urea (Mirsky and Pauling 1936), this article will concentrate on the quantitative analysis of heat-induced coagulation or gelation of egg-albumen proteins (Table 1). Factors affecting the kinetic parameters of gelation and the rheological properties of the gel will be discussed, as well as methods used to measure gelation.
a Adapted from Osuga and Feeney (1977) and Powrie
(1977); data refer to albumen from chicken eggs
b From Yang (1961)
c From Kuntz and Kauzmann (1974)
d From Vadehra and Nath (1973); ovomucoid pH 3.8-4.6;
ovomacroglobulin: pH 6-9
e From Fasman (1976)
A discussion of gelation necessitates defining some commonly used terms associated with the phenomenon. At first glance, some of these terms, may appear to overlap in meaning. However, each has its particular role in this discussion.
Attempts to describe the mechanism and theory of gel network formation are numerous. The classic explanation of heat induced aggregation of protein molecules is the following two-step process (Ferry, 1948):
Native protein -> denatured protein (long chains) -> aggregated protein (associated network)
The first step is considered a denaturation process and the second step an aggregation process. Comparison of the rate of the denaturation step vs that of the aggregation step helps determine gel characteristics. For example, Ferry (1948) suggested that for a given rate of denaturation the rate of aggregation will be slow if the attractive forces between the denatured protein chain are small. The resulting gel will be a finer network of protein chains, will be less opaque, and will exhibit less syneresis than one with a faster rate of aggregation. A coarser network of protein chains yields an opaque gel with large interstices capable of holding solvent which is easily expressed from the matrix. Hermansson (1979) suggested that conditions favoring denaturation, such as high or low pH, have the opposite effect on aggregation of globular proteins, possibly due to the fact that at high net charge, protein-solvent interactions such as denaturation are favored, rather than protein-protein interactions such as aggregation. A gel network with a certain degree of order can be attained if the aggregation step occurs more slowly than the denaturation step. thus giving the denatured protein molecule, time to orient themselves before aggregation; this is lower in opacity and higher in elasticity then one where aggregation is not suppressed. Schmidt (1981) suggested that if aggregation occurs simultaneously with denaturation, an opaque. less elastic gel results.
Since the kinetics of the denaturation step relative to the aggregation step appear to he important in determining the type of gel produced, it is useful to review some kinetic terms that aid in describing the gelation process:
l. Reaction Rate Constant. The first is the reaction rate constant k (min -1), which is obtained from the first-order relationship (Land, 1975):
dc
- -- = kc
dt
where c=concentration and -dc/dt=the rate at which concentration decreases. Integrating between limits et at time l=U and c et time t gives:
A plot of ln c vs t gives a line of slope -k.
The rate constant may be temperature dependent, end this dependence of reaction rate constant on temperature can best be described by the Arrhenius equation:
k=S exp (-Ea/RT)
where s=frequency factor (min -1), e.=activation energy (cal/mole). r=gas constant (1.987 cal/mole.°K), and t=absolute temperature (°K). A plot of In k vs 1/T gives e straight line of slope -Ea/R.
2. Z Value. Another useful kinetic quantity in describing the temperature dependency of the reaction rate is the Z value, which a defined as the necessary rise in heating temperature (°C) needed for a 10-fold increase in the reaction rate (Dagerskod, 1977). If some parameter of the gelation process is measured, say heating time to reach a certain gel strength, then the Zc value is the temperature rise needed to increase the heating time or reaction rate 10-fold, This value is useful in calculating the cook value, which is the time required at temperature T for a sample with a certain Zc value to receive a heat treatment equivalent to 1 min at 100°C (Dagerskod, 1977). To quantitate gelation, it in useful to coagulate samples to equal cook values to ensure equivalent heat treatment
Many researchers have tried to identify the factors which effect aggregation of proteins, since altering the rate of aggregation relative to the rate of denaturation appears to affect gel characteristics. The following are some of the factors which have been studied:
Shimada end Matsushita (1980a; 1981) found that the turbidity of
a 4.5% protein solution (3.6% ovalbumin, 0.9% conalbumin) heated
for 15 min at 80°C decreases as the pH increases from 8 to 11,
after which no turbidity is observed. Hardness of the gel is a
maximum at pH 8.5 and decreases on either side of this pH. The
turbidity increases upon addition of cations (Ca++
>> Li+ > Na+  Cs+) to a 2.8% albumin solution heated for 30 min at
80°C. Anions increase the turbidity in the alkaline pH regions
(SO4= > Cl- > Br-
> I- > SCN-). Salts shift the critical pH for
coagulant formation to alkaline pH levels as ionic strength
increases, perhaps by decreasing the amount of water bound to the
albumin and making it less soluble (Bull and Breese, 1970) or by
decreasing water protein interactions and increasing protein-protein
interactions.
Cs+) to a 2.8% albumin solution heated for 30 min at
80°C. Anions increase the turbidity in the alkaline pH regions
(SO4= > Cl- > Br-
> I- > SCN-). Salts shift the critical pH for
coagulant formation to alkaline pH levels as ionic strength
increases, perhaps by decreasing the amount of water bound to the
albumin and making it less soluble (Bull and Breese, 1970) or by
decreasing water protein interactions and increasing protein-protein
interactions.
Holme (1963) observed that the rate of aggregation of heated l.8% ovalbumin solutions is higher at pH 5.5 and 8.5 then at pH 7.0. However, at pH 5.5 the development of turbidity is rapid, with actual precipitation of the protein and slight increases in the levorotation and intrinsic viscosity of the supernatant, while at pH 8.5 there is slight turbidity and marked increases in levorotation and viscosity of the clarified solution.
Hegg et al. (1979) found from differential scanning calorimetry measurements that the denaturation temperature for a 4.4% ovalbumin solution increases from 62°C at pH 3 to 79°C at pH 6-10 but decreases to 77°C al pH 11. The aggregation temperature, defined as the temperature at which 10% of the protein has aggregated, was found to be 65-70°C at pH 4.0-5.5. The fact that aggregation only occurs around the isoelectric point implies that aggregation behavior is determined by the number of net charges on the ovalbumin molecule. When 1% NaCl is added to the ovalbumin, aggregation occurs at conditions ranging from 53°C at pH 3 to over 85°C at pH 11. The relationship between denaturation temperature TD and aggregation temperature TA determines the quality of the thermal aggregates; if TA is 6 -7°C lower than TD, precipitates of over 13% dry matter result; if TA is 4-6°C lower than TD, gel-like precipitates of 9-13% dry matter appear: if TD-TA is 2-4°C, opaque gels of 5-9 % dry matter form; and if TD-TA is 1-2°C or negative, transparent gels of less than 5% dry matter are found. Unlike Ferry (1948), Hegg et al. (1979) implied that aggregation takes place prior to denaturation, at all pH levels except pH 11, but that TD-TA decreases as pH is increased. At pH 10.5, denaturation and aggregation are predicted to occur simultaneously. Hegg and Lofqvist (1974) hypothesized that at extreme pH values or upon addition of anionic detergents the greater number of charged groups of the same sign repel each other and cause the protein to expend. The expanded molecules also repel each ether and thus hinder aggregation.
When liquid egg white is heated at 58°C for 5, 15, 30, and 60 min, the optical density decreases as the pH increases from 6.0 to 10.0 (Seideman at al., 1963); this agrees with analogous work using dilute albumin solutions. Egg white containing 50% sucrose requires a higher temperature for coagulation, which is also pH dependent.
Shimada and Matsushita (1980a; b) observed that as albumin concentration increases from 0.09%, to 12.5%, turbidity of the solutions heated at 80°C for 15 min is observed over a wider pH range. The pH required for equal turbidity increases as the protein concentration increases on the alkaline side of the isoelectric point from pH 6 to 11, and decreases on the acidic side from pH 4.5 to 3.
Many methods for measuring gelation involve testing individual samples after discrete time intervals of heating until coagulation or gelation is completed. Some of the methods which have been used are discussed below:
Using a capillary extrusion apparatus fitted to an Instron machine, Gossett (1983) and Gossett et al. (1983a) measured the apparent viscosity and yield force of albumen gels at various pH levels and heating temperatures; they found that gel strength increases with increasing pH and with increasing heating temperature (Table 2).
%DM=C0V0a/Wa
where, C0 =initial protein concentration, V0 =initial solution volume, a= percent aggregation and Wa = weight of the aggregate. Nakamura et a1. (1978) have measured the amount of aggregated proteins as the difference between the soluble protein concentration before and after heat treatment and centrifugation.
Fig. I -Typical Plots of electrobalance force vs time of heating for egg albumen (PH 9.0) at various temperatures of hearing. Plots shown terminate at or shortly after maximum force values.
The above methods for measuring heat-induced coagulation do have certain limitations, one of which is the need to make single-point measurements on many samples at different time intervals throughout the process to obtain a continuous view of the phenomenon. These samples must be [cooled?] instantly; otherwise the coagulation process con[tinues?] another limitation of some gel-strength or gravimetric measurements is that the sample is destroyed during the measurement. Yet another limitation is that for some turbidity measurements, once the sample becomes opaque, no further change is registered by the spectrophotometer despite continuing changes in gel characteristics. Furthermore, a gel can be clear end still exhibit increasing gel strength with time.
With these limitations in mind, we devised a new method to quantitate, the gelation process (Gossett et al., (1983b). This nondestructive technique requires only one sample to observe the total process. It involves continuous monitoring of the force exerted by the gel as coagulation takes place and can be used to obtain kinetic data on the heat-induced gelation of egg albumen.
The technique uses a recording electrobalance from which a wire probe is suspended into a jacketed cylinder containing the albumen sample. The cylinder is connected to a water bath maintained at the appropriate temperature, and, as coagulation occurs with time, the force exerted by the gel on the probe as the matrix forms is recorded on e linear recorder.
Typical electrobalance force-vs-time curves (Fig. 1) for albumen heated at temperatures from 60 to 95°C show that the force increases until a maximum force is observed. For some treatments, however, there are decreases in force after the maximum force is attained; this is thought to be due to expansion of the gel once the gel coagulates—the expansion may have raised the probe and caused a decrease in observed force.
a Quadruplicate determinations. From Gossett )1983),
Gossett et al. (1983b)
b Time at which first step "ends" and second step
"starts"
defg Means in the same column within a set that are
followed by different superscripts are significantly different (P
< 0.05)
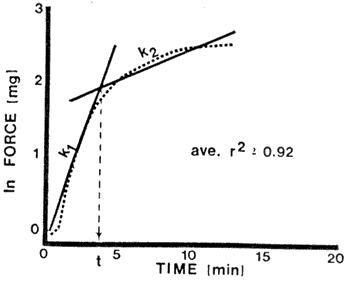
Fig. 2-PLOT OF ln FORCE VS TIME showing rate constants k1 and k2 and the time t where the first step "stops" and the second step "starts." From Gossett at al. (1983b)
Rate constants for albumen (at pH 9) heated at 60-9.5°C and for albumen (pH 7.0-11.0) heated at 80°C are shown in Table 3. First-order kinetics and the existence of two reactions during the heat-coagulation of the albumen, as su[illegible] by the broken-line curve in Figure 2, yielded rate [constants] k1 and k2 and were obtained by best r2 fit If k1 is interpreted to be the rate constant for the denaturation process and k2 the rate constant for the aggregation process, then the data suggest that the denaturation end aggregation rates both increase with increasing heating temperatures. The time at which the first step "ends" and the second step "starts" is found to decrease with increasing heating temperature; this suggests that at higher temperatures, denaturation occurs faster and the onset of aggregation is expedited. With increase in pH, rate constants for denaturation increase but those for aggregation decrease; the results agree with those of Nakamura et al. (1978) and Hermansson (1979), who suggested that aggregation is decelerated at higher pH levels.
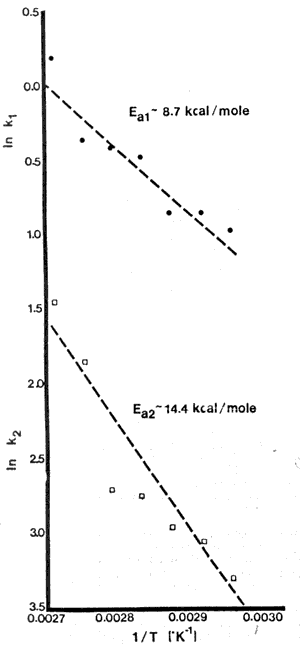
Fig 3-ARRHENIUS PLOTS for rate constants k1 (r=0.94) and k2 r=0.931) From Gossett et al (1983b)
Activation energies for the rate constants were calculated using the Arrhenius relationship; Figure 3 shows Ea to be 8.7 kcal/mole for the denaturation step and 14.4 kcal/mole for the aggregation step (Dwek and Navon, 1972, estimated Ea for the denaturation of egg albumen to be approximately 24 kcal/mole). In addition, a Zc value of 41.7°C was calculated by plotting the ln of the heating time required to reach a certain force value vs the heating temperatures.
Although this method offers the advantage of using just one a ample to continuously observe the gelation process, there are presently some limitations to the technique. The true source and physical meaning of the measured force are not yet understood and should be a topic of future research. Change. in the geometry of the gelling apparatus need to be explored to differentiate between the contribution of gelation and that of thermal expansion to the force. Lastly, optimization of probe design for various gel types would be desirable.
The gelation of food proteins is important to textural and rheological characteristics of a food product. Future, research should be directed toward finding quick, reliable methods that quantitate the gelation phenomenon so that food scientists can better manipulate the process to obtain desirable, high-quality foods.
Beveridge, T., Arntfield, S., Ko, S., and Chung, J.K.L. 1980. Firmness of heat-induced albumen coagulum. Poultry Sci. 59: 1229.
Bull, H.B. and Breese, K. 1970. Water and solute binding by proteins. I. Electrolytes. Arch. Biochem. Biophys. 137: 299.
Buttkus, H. 1974. On the nature of the chemical and physical bonds which contribute to some structural properties of protein foods: A hypothesis. J. Food Sci. 39: 484.
Cotterill, O.J. 1977. Freezing egg products. In "Egg Science and Technology," W.J. Stadelman and O.J. Cotterill, eds., 2nd ed., p. 145. AVI Pub. Co., Westport, CT.
Dagerskod, M. 1977. Time-temperature relationships in industrial cooking and frying. In "Physical, Chemical and Biological Changes in Food Caused by Thermal Processing," T. Hoyem and O. Kvale, eds., p. 77. Applied Science Publishers, Ltd., London.
Donovan, J.W., Mapes, C.J., Davis, J.G., and Garibaldi, J.A. 1975. A differential scanning calorimetric study of the stability of egg white to heat denaturation. J. Sci. Food Agric. 26: 73.
Dunkerley, J.A. and Hayes, J.F. 1980. Characterization of whey protein gels using a temperature gradient block. New Zealand J. Dairy Sci. Technol. 15:191.
Dwek, R.A. and Navon, G. 1972. On boiling an egg. Nature 240: 491.
Egelandsdal, B. 1980. Heat-induced gelling in solutions of ovalbumin. J. Food Sci. 45: 570.
Fasman, G.D. 1976. "Handbook of Biochemistry and Molecular Biology. Vol. II. Proteins," 3rd ed. CRC Press, Cleveland, OH.
Ferry, J.D. 1948. Protein gels. Adv. Prot. Chem. 4: 2.
Gossett, P.W. 1983. Some physical and rheological properties of raw and coagulated pH-adjusted or succinylated egg albumen with respect to water retention properties. Ph.D. thesis, Cornell Univ., Ithaca, NY.
Gossett, P.W. Rizvi, S.S.H., and Baker, R.C. 1983a. Selected rheological properties of pH-adjusted or succinylated egg albumen. J. Food Sci. 48: 1395.
Gossett, P.W., Rizvi, S.S.H., and Baker, R.C. 1983b. A new method to quantitate the coagulation process. J Food Sci. 48: 1400.
Haschemeyer, RH. and Haschemeyer, A.E.V. 1973. "Proteins," p. 353. John Wiley and Sons, NY.
Hegg, P.O. 1982. Conditions for the formation of heat induced gels of some globular food proteins. J. Food Sci. 47: 1241.
Hegg, P.O. and Lofqvist, B. 1974. The protective effect of small amounts of anionic detergents on the thermal aggregation of crude ovalbumin. J. Food Sci. 39: 1231.
Hegg. P.O., Martens, H., and Lofqvist, B. 1978. The protective effect of sodium dodecylsulphate on the thermal precipitation of conalbumin. A study on thermal aggregation and denaturation. J. Sci. Food Agric. 29: 245.
Hegg, P.O., Martens, H., and Lofqvist, B. 1979. Effects of pH and neutral salts on the formation and quality of thermal aggregates of ovalbumin. A study on thermal aggregation and denaturation. J. Sci. Food Agric. 30: 981.
Hermansson, A-M. 1979. Aggregation and denaturation involved in gel formation. In "Functionality and Protein Structure," A. Pour-El, ed., ACS Symp. Series 92, p. 81. Am. Chem. Soc., Washington, DC.
Hickson, D.W., Dill, C.W., Morgan, R.G., Sweat, V.E., Suter, D.A., and Carpenter, Z.L. 1982. Rheological properties of two heat-induced protein gels. J. Food Sci. 47: 783.
Holme, J. 1963. The thermal denaturation and aggregation of ovalbumin. J. Phys. Chem. 67: 782.
Johnson, TM. and Zabik, M.E. 1981. Gelation properties of albumen proteins, singly and in combination. Poultry Sci. 60: 2071.
Kuntz, ID. and Kauzmann, W. 1974. Hydration of proteins and polypeptides. Adv. Prot. Chem. 28: 239.
Lund, D.B. 1975, Heat processing. in "Principles of Food Science. Part II. Physical Principles of Food Preservation," O.R. Fennema, ed., p. 28. Marcel Dekker, Inc., NY.
Matsuda, T., Watanabe, K., and Sato, V. 1981a. Heat-induced aggregation of egg white proteins as studied by vertical flat-sheet polyacrylamide gel electrophoresis. J. Food Sci. 46: 1829.
Matsuda, T., Watanabe, K., and Sate, V. 1981b. Independent thermal unfolding of ovomucoid domains, Biochem. Biophys. Acta 669: 109.
Matsuda, T., Watanabe, K., and Sate, V. 1981c. Temperature-induced structural changes in chicken egg white ovomucoid. Agric. Biol. Chew. 45:1609.
Mirsky, A.E. and Pauling, L. 1936. On the structure of native, denatured, and coagulated proteins. Proc. Natl. Acad. Sci. 22: 439.
Nakamura, R., Sugiyama, H., and Sato, Y. 1978. Factors contributing to heat-induced aggregation of ovalbumin. Agric. Biol. Chem. 42: 819.
Osuga, D.T. and Feeney, RE. 1977. Egg proteins. In "Food Proteins," J.R. Whitaker and S.R. Tannenbaum, eds., p. 209. AVI Pub. Co., Inc., Westport, CT.
Powrie, W.D. 1977. Chemistry of eggs and egg products. In "Egg Science and Technology," W.J. Stadelman, and O.J. Cotterill, eds., 2nd ed., p. 65. AVI Pub. Co., Inc., Westport, CT.
Scheraga, HA. 1961. "Protein Structure," p. 81. Academic Press, New York.
Schmidt, RH. 1981. Gelation and coagulation. In "Protein Functionality in Foods," J.P. Cherry, ed., ACS Symp. Series 147, P. 131. Am. Chem. Soc., Washington, D.C.
Seideman, WE., Cotterill, O.J., and Funk, E.M. 1963. Factors affecting heat coagulation of egg white. Poultry Sci. 42: 406.
Shimada, K. and Matsushita, S. 1980a Thermal coagulation of egg albumin. J. Agric. Food Chem. 28: 409.
Shimada, K. and Matsushita, S. 1980b. Relationship between thermocoagulation of proteins and amino acid compositions. J. Agric. Food Chem. 28:413.
Shimada, K. and Matsushita, S. 1981. Effects of salts and denaturants on thermocoagulation of proteins. J. Agric. Food. Chem. 29: 15.
Vadehra, D.V. and Nath, KR., 1973. Eggs as a source of protein. CRC Crit. Rev. Food Technol., 4:193.
Yang, J.T. 1961. The viscosity of macromolecules in relation to molecular conformation. Adv, Prot. Chem. 16: 323.