Usage Recommendations for α-Amylases: Maximizing Enzyme Activity while Minimizing Enzyme-Artifact Binding Residues
by Harold M. EricksonManuscript available as PDF
Abstract
The results of spectroscopic investigations of reversible and irreversible enzyme binding to paper are reported, as well as the results of surface spectroscopic investigations of the extent to which such residues can be removed by rinsing. As much as 10% of the á-amylase present during a treatment under typical, non-stringent conditions may remain bound to a paper artifact after ethanol denaturation. Less than 30% of such denaturation residues were removed by subsequent washes with water, ethanol, or a water/ethanol 50% (v/v) mix. Alternative methods of inhibiting enzymes without affecting enzyme solubility are proposed, as are alternative methods of removing enzymes. Another common problem with enzyme usage is that overly strong concentrations and elevated temperatures are used to compensate for reduced effectiveness of the enzymes under indifferently chosen treatment conditions; criteria for selection of enzymes and adaption of treatment conditions are suggested. Animal, cereal, fungal and bacterial á-amylases are reviewed and fungal and bacterial á-amylases are recommended for various applications. Formulas and sample calculations for bath immersion, topical, and viscous media applications are presented. Use of acetate buffers rather than phosphate buffers is proposed.
Introduction
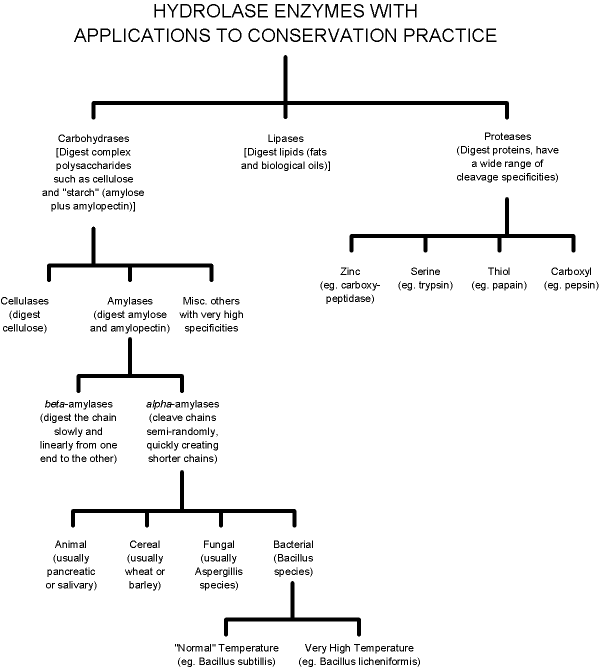
Figure 1. Partial taxonomy of hydrolases illustrating context of various α-amylases
The proteins capable of catalyzing physiologically important biochemical processes are known collectively as enzymes. Among these are a group whose function is catalyzing hydrolytic cleavage (digestion) of such biological polymers as proteins, starches and fats. These are known as hydrolases (see Figure 1). The focus of this paper will be the hydrolases whose function is the digestion of complex polysaccharides (carbohydrates) such as starch and cellulose. These enzymes, known as carbohydrates, are highly specific, each catalyzing the hydrolytic scission of a specific glycosidic bond. Amylases are carbohydrates that catalyze the digestion of the α-D-1,4 glycosidic bonds in the amylose and amylopectin that are the primary components of the purified starches used in making paste. The amylases effect a rapid reduction of the length of the starch polymer. The resulting fragments are oligosaccharides that are readily soluble in water and that are too short to retain significant adhesive capability.
These enzymes have been used in conservation for years, usually with scant effort expended to maximize their catalytic efficiency. In order to compensate for the amylases' reduced effectiveness, the concentrations used have often been higher than was necessary. Treatment of this sort has often been followed by a step in which the enzyme is "inactivated" by exposure to ethanol or hot water; the intent of this usually ineffective and potentially dangerous step is to disrupt the enzyme's tertiary shape, thereby terminating its hydrolytic capacity. Under treatment conditions, especially when ethanol inactivation is used, some fraction of the enzyme present may bind to a paper or textile artifact as a residue. It should be noted that the significance of such residues can be and has been debated, especially in light of the fact that such residues are essentially identical to those produced in much heavier quantities by gelatin-sizing. Nevertheless, since the implications—if any—of such enzyme residues are presently outside the realm of consensus, it is prudent to minimize the extent of such binding, at least to the extent permitted by other conservation and curatorial considerations.
This paper has two primary goals. The first is to report briefly the results of spectroscopic investigations of the nature and extent of protein binding to paper artifacts that occurs under typical treatment conditions, as well as spectroscopic investigations of the extent to which such residues can be removed by rinsing. The other objective is to present guidelines for amylase usage that are drawn from a review of industrial and patent literature. These protocols are designed to minimize such protein-artifact binding by maximizing the digestive effectiveness of the amylase. While this article focuses on applications to paper artifacts, the results are sufficiently general that they should also be of use in textile conservation.
Enzyme-Paper Interactions
A recent conservation treatment involving α-amylase and proteases prompted an investigation of the extent to which such enzymes would reversibly and/or irreversibly bind with the lignocellulosic structure of paper. A multiplexing ultraviolet/visible (UV-vis) spectrophotometer was used to monitor the adsorption of ethanol-denatured α-amylase to paper. This work was performed under conditions encountered within protocols still commonly used by the conservation community. Analysis revealed that 10% or more of the denatured α-amylase present may bind to the paper object being treated when denaturation of the enzyme occurs in the presence of the cellulosic object. Further investigation using a UV-vis spectrophotometer with surface absorption accessory revealed that rinsing protocols practiced in the conservation community typically remove 30% or less of the enzyme that becomes bound to the paper during these unnecessary denaturation steps. Details of these results may be found in the EXPERIMENTAL section of this paper.
Usage Considerations
The bench conservator's work with enzymes is always constrained by the professional consensus that treatment must never irreversibly change an artifact. Two important limitations on enzyme use are implicit in this ethic. The first is that enzyme residues such as those described above should be minimized or eliminated—at least until their consequences are fully understood. This is accomplished through thoughtful attention to selection of the enzyme and to its usage requirements. The other limitation is that treatment conditions when using enzymes must be governed primarily by concern for the integrity of the artifact, and must not be determined by usage optima of the enzymes. Given these constraints, many conservators are understandably hesitant to use these powerful biomolecular tools. While a healthy dose of such caution is desirable, it is also important that the conservator also be aware that judicious selection of a particular α-amylase type and purity, when made with commensurate anticipation of reaction conditions, can yield safe and effective treatment for the majority of objects. The discussion that follows is intended to prepare the conservator to match the artifact's frailties and susceptibilities to a particular α-amylase's reaction optima.
The conservation community has been repeatedly cautioned by Burgess and others not to use pH, temperature and ion concentration information from the assay parameters supplied with the assay results, usually on the container's label. These are standardized conditions that are used to assay any á-amylase, regardless of its source and its reaction optima. The intent of these standardized conditions is merely to permit the comparison of enzymes from various suppliers. Users should exercise similar caution about the use of reaction optima data from the biochemistry and molecular biology literature, if it is to be used at all, because that data is almost universally generated in studies in which both the enzyme and the digestible substrate are dissolved in aqueous solution with careful attention to ion balance and pH. The applications confronting the bench conservator, by comparison, are invariably bi-phasic. They take place at the interface between a solid substrate- typically a cross-linked adhesive- and a fully or partially solvated enzyme. Furthermore, the requirements of a particular conservation application seldom conveniently match the digestion optima of the enzyme sitting in the freezer down the hall.
Fortuitously, these same problems of reaction-phase heterogeneity and difficult reaction conditions also confront the brewing, corn syrup, cheese-making, candy, baking, dairy, meat, seafood processing, vegetable processing, starch, paper and textile industries. Dozens of volumes of data are available (2, 3, 4, 5, 6, 7, 8, 9, 10, 11) as are hundreds of technical papers and hundreds of patent applications. The half-billion dollar-per-year enzyme industry generates dozens of patent applications annually, each of which is supported by masses of such data. Many of these patent applications involve amylases and proteases.
I believe that the following summary of that body of industrial data largely complements and confirms forthcoming experimental results generated independently by investigators at the Canadian Conservation Institute. The industrial patent literature is rich in experimental data describing behavior of enzymes under real-world treatment conditions—the same non-ideal conditions that perplex the conservator. Among the conditions to which the practicing conservator must pay close attention are the pH, the calcium and sodium concentrations, the temperature, and the commercially available purity of the enzymes. Each of these factors varies widely and is dependent on the species from which the amylase was extracted. Conveniently, the industrial labs have already completed most of the investigations of these factors.
Figure 1 outlines some of the many available variants of amylases. One of the most important considerations is the choice of α-amylases over β-amylases. β-amylases are mechanistically exo—that is, they methodically digest the amylose and amylopectin into di- and tri-saccharides, working linearly from one end of the polymeric chain to the other. They suffer from the drawback that they are stymied when they reach a α-D-1,6 branching linkage in amylopectin, which occurs on average about once in every 25 linear glucose units, the remainder of which are joined by the more typical α-D-1,4 linkages. α-Amylases, which conversely are mechanistically endo, attack the starch polymeric structure semi-randomly, quickly reducing it to a series of readily-soluble short oligosaccharides. The closely related glucoamylases and pullulanases will be encountered in the amylase literature by any enquiring reader, but these are not well suited to conservation usage. Glucoamylases hydrolyze both α-D-1,4 and α-D-1,6 linkages, but are mechanistically exo, making them too inefficient and slow for conservation treatments. Pullulanases, on the other hand, are debranching enzymes that only digest the α-D-1,6 linkages that are responsible for amylopectin's branched structure; they therefore could be used as a complement to β-amylases, but β-amylase's slow exo digestion eliminates even the complementary pair from consideration as useful biomolecular tools.
α-Amylases are extracted from a number of biological sources. These sources include animal (usually pancreatic or salivary), cereal (usually wheat or barley) 12, fungal (usually derived from large-scale fermentations of Aspergillus species), and bacterial (derived from similar fermentations of Bacillus species).13 Even within a given species, amylase optima may vary depending on the organ from which or fermentation conditions under which the amylase was extracted. Selection is further complicated by the fact that industrial suppliers of enzymes have cultivated mutant strains of many popular amylase-producing species, each of which in turn has different digestion condition optima.
It will be seen that economics and availability of high purities will largely dictate the use of fungal and bacterial α-amylases. A detailed discussion is found in the Specific Usage Recommendations section.
Pre-Testing for Artifact Safety
In at least one conservation treatment whose results were examined in preparing for this paper, use of an enzyme had a substantial negative impact on the object that was treated. In that particular case, an immersion bath of protease resulted in a sudden floating of a number of mold-damaged fragments that were apparently being tenuously held to the bulk of the paper by the adhesive action of the gelatin size. While this is not a problem likely to occur in amylase treatments of Western paper objects, it points up the need for pre-testing, even with a biomolecular tool whose specificity is as high as that of amylase. It is especially imperative that workers involved with Islamic paper artifacts act with extreme caution, since these are known to be traditionally sized, burnished and even dyed using starch 14.
Some modern Western papers are starch coated or starch loaded. Preliminary investigations performed at the Bodleian Library showed no visible effects of amylase treatment on starch loaded or coated papers, or on the printing on such papers. The investigators did report some apparent but unquantified weakening of the paper 15. Stringent pretesting then is definitely merited when such Western papers are encountered.
There is good reason to believe that amylase activity would be statistically more likely to attack starch incorporated in the paper than to attack a starch-adhesive residue on the paper, based purely on the relative number of sites for digestive attack; amylase treatment is then probably contraindicated in both these cases because of the low probability of effective treatment and the significant possibility of ill effects.
Pretesting for Enzyme Activity
In an ideal world, conservators would be sufficiently comfortable with standard molecular biology techniques that they could perform their own assays of the hydrolytic activity of such enzymes as amylase and protease. Indeed, this has been recommended. Unsurprisingly, however, not one of the bench conservators interviewed in preparation for this paper was comfortable with the prospect of performing such an assay.
Given the excellent reliability of the enzyme market at present, particularly in light of the high purities and long shelf lives available, a few reasonable guidelines are probably sufficient to permit conservators to assume the acceptability of a given enzyme. The three guidelines are the following. Always purchase high purity enzymes, since crude preparations contain unnecessary contaminants, and may even contain such rogue enzymes as cellulase. Always maintain high standards of chemical practice when micropipetting from the bulk container or when transferring lyophilized solids to the scale, in order to minimize contamination of either the bulk product or the prepared solution; cross-contamination with protease must be avoided with particular diligence. Finally, one should always store enzymes as recommended by the supplier or as described in a later section.
If a conservator desires a crude but effective test for activity, then s/he may wish to adopt some form of the following test 16. Select two sheets of a strong uncoated paper whose furnish is neither extremely smooth nor extremely rough. Working quickly, brush out a fairly heavy coat of starch paste onto each, using paste that has been thinned to about half of its normal viscosity. Place the paste sides together and allow to dry completely (one to two days) under pressure. Place the dried composite sheet into a humid oven at 90-98°C (194-208°F) for 3-7 days until the adhesive is sufficiently cross-linked to be considered intractable 17. Cut the sheet into small strips, each pair of which will permit the checking of an amylase preparation once. Since this "assay" should only be necessary on those rare occasions when long-stored enzymes are removed from cold storage, a single preparation of such a set of strips should provide enough material to supply a number of labs for years.
The "assay" is performed by simply moistening one strip with an appropriately prepared enzyme solution and moistening the other with a control solution differing from the enzyme solution only in the absence of á -amylase. The pasted halves of the strip moistened with the enzyme solution should separate significantly more quickly than those of the strip moistened with the control solution.
Methods of Application
While the spectroscopic studies detailed in the EXPERIMENTAL section are focused on bath immersion, it should be noted that the implications generalize well to all other common methods of enzyme application. These methods include, but are not limited to, bath immersion (aqueous as well as partially or completely nonaqueous), topical or spot application with swabs or blotter paper, and viscous media (gel or poultice) application.
In bath immersions, a relatively large volume of dilute enzyme solution is used. The advantages of this method are that the enzymes are freely mobile, permitting more facile digestion of the starch substrate, and that the resulting oligosaccharide fragments dissolve away from the site of the digestion. The disadvantage, of course, is the incompatibility of many media with water. This incompatibility has been overcome in some treatments through the use of nonaqueous or partially aqueous solutions 18. The use of carbohydrates in nonaqueous media require overcoming some particularly difficult biophysical constraints 19.
In general, aqueous immersion bath treatments performed with adequate attention to ionic concentrations and pH should work with acceptable speed at concentrations of 1-5 units activity per milliliter 20 (see sample calculations, later section), but a number of other factors should be weighed in choosing the enzyme concentration to be used in a particular treatment. The dynamic that should be foremost in the conservators mind is the tradeoff between, on the one hand, the lengthened digestive time required at sub-optimal pH and temperature which permits more time for such undesirable effects as offsetting, and on the other, the potential damage to the object that may result from attempting to achieve shortened digestion times with elevated temperature and pH adjustment.
A good rule of thumb for α-amylase concentrations is viscous media and in solutions intended for topical application appears to be a minimum tenfold increase in enzyme concentration over what would be used for aqueous immersion. It is imperative to remember that a dry immobilized enzyme cannot perform any useful function. When performing such viscous media or topical applications, sufficient moisture must be maintained to yield mobility for the enzyme. Generally speaking, this will be the point at which the paper fibers have just become fully swollen but do not contain bulk water. This condition can be achieved in viscous media applications by using a loose gel or by slight prewetting; humidity is then maintained by the gel or poultice. Such a fiber-swollen condition can be maintained for topical applications by damp blotters, with or without a protective barrier of Gore-Tex® expanded poly(tetrafluoroethylene) on polyester backing.
The consensus among book-and-paper conservators appears to be that topical application is most common, followed by aqueous immersion. Other methods are rarely if ever used.
Temperature
The conservation literature frequently mentions the need to let enzymes work at elevated temperatures, as well as of the enzymes' tendency to denature at about 40°C. In fact, the amylases and proteases are quite robust thermally, with most reaching their thermal optima in the range of 50-105°C and losing significant catalytic activity in the range of 65-110°C. These figures make two critically important points immediately clear. First, attempts at denaturation with hot water would be ineffective at the conventionally recommended temperatures, and would be dangerous to the artifact at temperatures where effective denaturation is achieved. Second, temperatures sufficiently elevated to maximize the enzymes' catalytic activity are likely to be dangerous to the object.
None of this should be taken to belittle the intended result of working at elevated temperatures; indeed, digestion rates can be increased hundred-fold in this way. What the bench conservator should consider, however, is that similar increases can be achieved through careful selection of enzymes and through careful attention to pH and ion balance, while eliminating the logistical difficulties and artifact jeopardy that result from attempts to work at these temperatures.
Also, those conservators who perform topical application of "warm" enzyme solutions by dipping swabs or blotter paper in a beaker of enzyme solution that is kept at the benchtop on an electric hotplate are accomplishing little if any benefit for their extra trouble. The solution will have largely cooled by the time it is transferred to the object, and the digestion temperature will be determined by the temperature of the object. In fact, such a procedure may have a negative impact since the enzyme will suffer accelerated deterioration while sitting for hours on the benchtop.
pH Control and Buffer Solutions
While it is hoped that the guidelines from the SPECIFIC USAGE RECOMMENDATIONS section will be adequate to allow the conservator to match an appropriate enzyme to the treated object's native pH, there may be times when the pH of the enzyme solution or poultice must be controlled in order to maintain the activity of the enzyme. Phosphate buffers have traditionally been used in conservation practice to control enzyme treatment pH. Phosphates, however, have the unfortunate side effect of causing precipitation of calcium ions that are needed for full activity of the fungal and bacterial amylases. Since these microbiological amylases are economically and chemically best-suited to conservation treatment, this calcium phosphate precipitation presents a significant problem.
A good solution would be to use acetate buffers instead of the phosphate buffers. Data from Wakim, et al, can be used to demonstrate that no significant change in activity occurs when buffer systems are changed, at least for porcine pancreatic á-amylase,21which is not so highly calcium dependent.
The acetate buffers are simple to prepare. 0.1 ml of concentrated acetic acid ("glacial," 99.4-99.8% CH3COOH, density 1.05g/ml) combined with 1.44 g sodium acetate (NaC2H3O2) or 2.38 g sodium acetate trihydrate (NaC2H3O2 . 3H2O) in a liter of solution will yield a simple buffer with a pH of about 5.75. This simple buffer displays only limited resistance to pH change by strong bases, but it does exhibit excellent resistance to pH changes by acids, which is a far more prevalent complication in conservation applications anyway. pH of the buffer can be further increased to a value as high as ph=8.5 by further decreasing the volume of glacial acetic acid added to the buffer, although at the cost of still more limited capacity to respond to an object having a native pH greater than 8.5. The prime advantage of acetate buffers is that they do not interfere with the solubility of any metallic ions, while also having no known conservation contraindications. Of particular comfort is the fact that acetates are so soluble that they rinse easily from the paper.
A note in passing about Trizma®. buffers is in order. Such buffers have been recommended to avoid the phosphate precipitation problem described above, while providing full buffering against both acids and bases. Unpublished results indicate that Trizma®. buffers may decrease fold endurance of paper. Until this issue is resolved, acetate buffers appear to be the preferable alternative.
Most importantly, since the majority of treatments are apparently topical rather than immersion, the conservator should keep in mind that buffering of an enzyme solution that is to be used for topical application is largely a futile endeavor, since the pH of the digestive process will be determined by the native pH of the object.
Ion Balance
á-Amylases are considered, broadly speaking, metallo-enzymes, i.e. enzymes that require the presence of a metal cation "cofactor" in order to express their full activity. In á-amylases, the metal is calcium. The number of calcium atoms required per enzyme have been reviewed in many places over the last 3 decades. The many apparent contradictions in this body of data are the result of an evolving awareness of the tendency of many enzymes, amylases among them, to form multi-enzyme quaternary complexes that may on first investigation appear to be larger enzymes having higher activity and higher calcium counts. Such issues may now be resolved unambiguously using x-ray diffraction.
Some á-amylases, notably those derived from mammalian sources, may also require the presence of other enzymes such as sodium and calcium. The requirements of various á-amylases are discussed in more detail in the SPECIFIC USAGE RECOMMENDATIONS section.
The presence of appropriate ions can easily increase the activity of the enzyme ten-fold or more, allowing the conservator to either lower the enzyme's concentration to minimize residual binding or to shorten the working time and thereby lower the likelihood of such aqueous treatment artifacts as offsetting of media or tidelines. The conservator, however, should not begin to dread the preparation of millimolar solutions of various ions in order to tweak the performance of a particular enzyme. The fungal and bacterial enzymes are quite robust; the requisite Ca+2 ion in Asp. species, for instance, appears to be so tightly bound that it will not be lost even if dissolved in distilled water. Furthermore, Toda and Narita have established that magnesium ions will restore the activity of Asp. oryzae α-amylases that have had their calcium removed by incubation with the calcium-chelator ethylenediaminetetraacetic acid (EDTA). 22
Concentration Calculations
The most commonly purchased form of enzymes is a water-soluble Iyophilized powder. The number of grams of enzyme that will be weighed out, m,, when preparing V milliliters of an enzyme solution having a concentration of C, units activity per ml solution is given by:
Equation 1
where A, is the activity of the solid expressed as "units of activity per mg of solid" (as opposed to "units of activity per mg of protein"). If, for instance, one needed to make 100 ml of an enzyme solution having an activity concentration of 5 units/ml from a powder labeled "1100 units/mg solid," then one would weigh out:
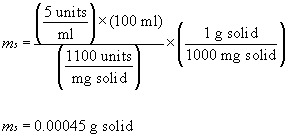
Equation 2
It will be obvious that the measurement of such small masses will require an analytical scale having a minimum sensitivity of 0.0001 g and ideally 0.00001 g. Many labs, having access only to scale less sensitive than this, may need to adopt the expedient of making larger quantities of solution and storing the excess, or of making a concentrated stock solution that can be stored in frozen aliquots.
Some enzymes, however, are readily available only in a concentrated liquid form. One purchases a large amount of highly purified enzyme, say 500,000 units or more, in a volume of 30-500 ml of aqueous buffer solution. This actually makes the preparation of solutions easier, as long as one has access to a micropipette capable of measuring down to at least ten microliters (10 p1). These micropipettes are available affordably from a number of sources and should be purchased with a supply of tips that are ready-packed in their dispenser boxes.23The calculation of the number of μl of concentrate, Vμ, that should be dispensed when preparing V milliliters of an enzyme solution having a concentration of C, units activity per ml solution is given by:
Equation 3
where ABE is the number of units of activity of bulk enzyme that were purchased, and where VBE is the approximate volume of the bulk enzyme concentrate that was purchased. If one had purchased, let us say, 500,000 units of B. licheniformis which arrived dissolved in approximately 35 ml of bulk solution, 24 and wished to prepare 10 ml of a treatment bath having a concentration of 15 units/ml, then one would measure out :
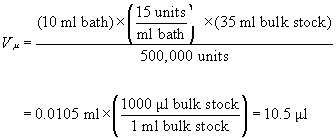
Equation 4
Storage of Enzymes
Amylases and proteases are notoriously robust enzymes. The conventional wisdom is that if one smeared saliva into a dirty table and left it to dry fully exposed for weeks, one could still find significant retention of catalytic activity in an extract of scrapings from the table. That said, some qualifiers are in order. Enzymes are traditionally stored at temperatures just above or below the freezing point of water in order to preserve their activity. Users must keep clearly in mind that freeze-thaw cycles are one of the worst culprits in accelerated loss of enzyme function. Concentrated bulk stock solutions are typically stored refrigerated at 0-5°C in order to avoid these cycles. Premeasured aliquots of concentrated stock solutions, or even premixed ready-to-use solutions, may be stored frozen for six or even twelve months, but only in a freezer that does not have an automatic defrost cycle. The only freeze-thaw to which these should be exposed is the initial freeze and final thaw. Ready-to-use solutions may be stored refrigerated at 0-5°C for weeks, but users should bear in mind that an additional factor then enters play. Not only are traditional modes of inactivation slowly taking their toll, but precipitation of the protein on the container wall will also be contributing more rapidly to loss of solution activity. For this reason, Sigma Chemicals recommends that only solutions with concentrations greater than 0.5 mg protein/ml solution be stored refrigerated. 25 Since these concentrations are significantly higher than those typically used in conservation practice, this method of storage is recommended only for premeasured aliquots of concentrated stock solutions.26Bulk enzymes received as lyophilized solids should be stored frozen in a desiccator, or at least inside multiple plastic bags with a desiccant. Properly stored bulk amylases, whether solid or concentrated liquid, should lose activity only at the rate of 1-2% per year.
In summary, then, amylase solids and concentrated liquid solutions are sufficiently robust to permit long term storage in the freezer and refrigerator respectively. The two storage problems that may arise are in the freezing of liquids in an auto-defrost freezer, and the refrigeration of dilute (working strength) solutions that will lose strength as enzyme precipitates on the glass walls of the container.
Deactivation of Enzymes
An enzyme's activity can be disrupted through any chemical, thermal or physical method that alters the tertiary configuration of the protein. The three broad classes of inactivation are inhibition, denaturation and destruction. Inhibition is usually accomplished by a change in pH or ion balance, or by the introduction of enzyme-specific inhibition proteins; it results in a reversible or irreversible blockage of access to and/or shape-change of the enzyme's active site. Denaturation—a partial or complete unraveling of the protein's tertiary structure—is usually accomplished through extreme pH or heat, simple drying, solvents (such as ethanol or acetone), or surfactants (ea. sodium dodecyl sulfate, SDS); the denaturation may be partially or fully reversible if the denaturant is removed or neutralized and if the denaturation is not too far progressed. Irreversible destruction involves chemically changing the nature of the protein; in conservation this would typically be achieved by permitting the air to oxidize a dried protein residue or by cleavage with a protease.
The conservation literature has conventionally prescribed the denaturation of enzymes with heated water or ethanol. These steps are unnecessary for several reasons. Foremost among these is the potential damage to the object that may result from the use of such harsh denaturation conditions, a consideration that appears in the literature at least as early as 1977.27 Supporting this contention is the fact that the water soluble enzymes will largely wash away if they are never permitted to dry or denature on the paper, and that any residues that do dry (as opposed to denature) on the paper will wash away after brief prehydration. Further supporting this contention is the fact that there exist no known negative effects of a dried enzyme remaining on the paper after, say, a topical spot application to a water-sensitive object, other than the difficulty of getting a starch paste to adhere to that spot in the near future.
Specific Usage Recommendations
When purchasing enzymes, avoid those described as crude preparations, since these will be contaminated with other proteins that bind to the artifact without conveying any additional digestive activity to the treatment solution. Indeed, crude preparations of this type may even be contaminated with cellulase that will inflict significant structural damage on the paper by hydrolytic digestion of the paper's cellulosic fibers. Purchase only high purity enzymes, usually described in catalogs as crystalline or lyophilized.
Burgess has recommended that enzyme concentrations for immersion treatment of paper objects be on the order of 1-5 units of amylase activity per milliliter of solution. Her rationale is that these concentrations are adequate to digest most occurrences of cross-linked starch adhesives encountered in conservation practice if careful attention is paid to digestion conditions such as pH, ion balance, and temperature.28 Remember, however, that these recommendations are for optimized conditions. Little if any detectable residue is likely to be deposited on the object even at concentrations in the range of 50-250 units activity per ml, as long as in situ denaturation is avoided. If an adhesive is intractable, do not hesitate to increase the á-amylase concentration significantly.
The paragraphs that follow address the issues pertinent to reaction conditions for the á-amylases of fungal, bacterial, cereal and mammalian origin respectively.
The most readily available fungal α-amylases are those derived from Aspergillus oryzae. These are available commercially in reasonably high purity and display the calcium inactivation behavior described below, making it a particularly good choice for risky treatments. The amylase derived from native A. oryzae displays acceptable activity between pH 5 and 7 with optimum activity reportedly between 4.8-5.8. Temperature stability extends to 50-55°C. It is important to note that fungal amylases require calcium ions for full enzymatic activity; this must not, however, be construed as a requirement for the addition of calcium ions to the treatment bath. The calcium ion needed to maintain the hydrolytic activity of fungal á-amylases is tightly bound in the active site of the enzyme protein 29, and the very small concentration of calcium leached from the artifact should be more than sufficient to maintain the enzyme's full activity. Fortification of the treatment bath with calcium ions is likely to inactivate the enzyme as described in a later section. The á-amylase extracted from cultures of Asp. niger has similar properties, but is not readily available in the purities required for conservation practice.
Bacterial amylases have been extensively researched and are prolifically described in the patent literature. Unfortunately, only two Bacillus enzymes are readily available to the conservation community in acceptable purity. Both B. subtillis and B. licheniformis require sodium ion and calcium ion in order to preserve the enzyme's full activity. That of B. licheniformis, however, requires only 5 ppm Ca+2, while that of B. subtillis requires 150 ppm Ca+2. The heat stability of B. subtillis amylase extends as high as 80-85°C and that of B. licheniformis extends to 110°C; both species' amylases nevertheless show comparable activities at room temperature.
Cereal α-amylases, such as those derived from barley malt or wheat are available only in crude form, which makes these enzymes unsuitable for most conservators. For conservators with access to preparative-scale protein chromatographic apparatus, these crude preparations do, however, offer an economical source of purifiable amylase. The user should be aware that cereal amylases display a pronounced shift in pH optima with change in temperature. For instance, the pH range of optimum activity of amylase derived from barley malt will shift from pH 4.7-5.4 at 50-55°C to pH 5.6-5.8 at 70-75°C. The conventional rule of thumb is that cereal α-amylases display their highest activity between pH 5-6 30.
Mammalian α-amylases, such as those extracted from beef or porcine pancreas or salivary gland, or indeed those from human saliva, are available in very high purity, but are prohibitively expensive, without offering additional advantages. Their use is further complicated by the need to supply chloride ions in order to achieve full enzymatic activity. While their optimal activity is traditionally said to be around pH 7, this activity can decline sharply and show a substantial loss of breadth of pH activity when chloride ion is absent. Porcine pancreatic α-amylase, for instance, has been shown to decrease activity by an order of magnitude when Cl--enrichment of 0.025 M is not provided, while simultaneously decreasing its range of acceptable activity from pH 5-10 to pH 5-6 31.
For conservation purposes, then, the issues of function, purity and economics alone are sufficient to limit the choice of available carbohydrases to fungal and bacterial α-amylases. The currently available options are α-amylases from B. subtillis, B. licheniformis, and Asp. oryzae, of which the most flexible, easy-to-dispense, and rapid working is B. licheniformis. Conservators looking for a single multi-purpose enzyme will do well to consider purchasing a micropipette and beginning to use B. licheniformis α-amylase.
Urgent Arrest of Aberrant Treatment
Recent research 32 indicates that the activity of α-amylases derived from such fungal species as Aspergillus oryzae can be effectively terminated using calcium ions -a reversible inhibitor. These investigators found that calcium concentrations of 20 millimolar (800 ppm Ca+2) were sufficient to reduce the enzymatic activity of fungal amylases by 99%. This offers the conservator who is attempting a high-risk immersion treatment the opportunity to arrest the treatment's progress in the event of undesirable effects. The conservator can prepare a 2 molar Ca+2 stock solution, premeasured in a volume equal to one-hundredth of the volume of the treatment bath; a 2 M Ca+2 solution may be prepared by dissolving 2.22 g calcium chloride, CaCl2, or 2.94 g calcium chloride monohydrate, CaCl2 -H2O, in 10 ml of distilled water. This solution is extremely stable and can be kept for future use in a tightly sealed container. A 10 ml aliquot should be prepared and ready for every 1 L volume of the bath. The conservator is then prepared to quickly inactivate the fungal α-amylase by pouring the calcium stock solution into the treatment bath with gentle agitation. The artifact may now be removed carefully from the bath, taking time to avoid physical damage, with the knowledge that further amylase-inflicted damage will not occur while the time is being taken to exercise this caution.
Environmental Health and Safety Issues
Enzymes present very few dangers to the user other than as an inhaled irritant and potential inhaled allergen. Skin absorption or ingestion are unlikely to occur and present only low hazard if they should occur. Keep clearly in mind that amylase and protease are two of the primary active ingredients of saliva.
Standard industrial hygiene precautions for handling low hazard friable, air-suspendable, powdered solids should be taken when transferring and weighing solid enzyme preparations. Standard precautions would include the wearing of an appropriately fitted respirator by any user with a known or suspected allergy to amylases. The suppliers' Material Safety Data Sheets (MSDS) may be consulted for further details, but the user should keep in mind that MSDS's for proteins such as enzymes tend to be filled with generic boilerplate text. The hazards of individual proteins cannot be adequately investigated for the thousands of commercially available proteins, each of which is sold only in very small quantities. This results in MSDS's that tend to be written for the worst possible case, whether or not the available evidence supports the recommended levels of industrial hygiene.
Spills of either the solutions or solids present no great hazards other than the minor health considerations described above. Solutions of amylase are roughly as dangerous as saliva and may be disposed of in approximately the same manner. Large quantities of unused solids should be disposed of through an appropriate hazardous materials disposal program, while small amounts of spilled or unused amylase can be safely wrapped in damp paper towels and thrown away. These practical guidelines may be superseded by legal requirements in communities, states or countries with broad and stringent regulations covering disposal of "chemicals" in the sanitary sewer or municipal landfills.
Experimental
A Hewlett-Packard HP8450A multiplexing UV-vis spectrophotometer was used to monitor the adsorption of ethanol-denatured amylase to paper. This work was performed using a variety of standard and historical papers under conditions encountered within protocols still in common use by the conservation community. An 0.1% (w/v) solution of amylase was prepared. The amylase used was Sigma Catalog No. A 6380, Type IIA, which was received as a four-fold recrystallized solid having 1400 units of activity per mg of solid 33; the enzyme had been supplied to Sigma as being of B. subtillis origin, but Sigma included a disclaimer in the catalog suggesting that their investigation indicated that the source would more properly be described as B. amyloliquifaciens.
The results indicate that a significant percentage of the denatured amylase will bind to a paper sample that is present at the time of denaturation. This effect is absent when paper is added to a solution of native enzyme and is only minor when the paper is added after denaturation, indicating that the enzyme-artifact binding primarily takes place immediately after denaturation, before partial renaturation can occur. More detailed data on the extent of the enzyme-artifact binding to a variety of historic and modern papers was sought, but the extent of UV-absorbing leachate from most papers was of such magnitude and variability as to overwhelm and obscure the relatively small spectral changes being monitored. Whatman #3 Chromatography (W3C) paper (Cat. CP3MM, basis weight 185 g/m2, thickness 0.33 mm), however, was shown to have only a negligible UV-absorbing leachate. This permitted observation of a quantitatively reproducible effect. When 2 ml of standard 0.1% (w/v) amylase aqueous solution 34, is denatured with 1 ml of ethanol in the presence of 6.25 cm2 of W3C paper, 11.6 + 0.3 % of the 2 mg of amylase present were bound to the paper. These results indicate that in situ denaturation is undesirable under almost all circumstances.
When deactivation of the enzyme can be avoided—which should be in virtually every case—simple rinsing will remove all measurable levels of enzyme from the paper.
Further investigation using a Perkin-Elmer Lambda 3B W-vis spectrophotometer with surface absorption accessory reveals that rinsing protocols typical of those practiced in the conservation community remove 30% or less of the enzyme bound to the paper. Generally speaking, it was found that two-minute washes with straight ethanol were consistently the least effective at removing amylase residues left after in situ denaturation, removing only 18-20% of the denatured protein residues adhered to the paper. Similarly, two-minute washes with 50% aqueous ethanol (v/v) removed 18-29% and with distilled water removed 27-30%. In each case, the 1 cm x 2 cm Whatman #1 Chromatography (W1C) paper samples rinsed were prepared by a five-minute soak in an 0.1% (w/v) α-amylase solution. This was followed by a two-minute in situ denaturation by addition of 50% (v/v) ethanol, finishing with a double blotting. The strips were then rinsed with gentle agitation in 10 ml of the above described wash baths. The strips were again double blotted, and were permitted to air dry. They were then subjected to surface UV-vis absorption spectrophotometry. Kubelka-Munk transformation of the resulting spectra was regarded as unnecessary because of the thinness of the W1C paper.
The Bottom Line
- Treatment conditions must be dictated by the needs of the object rather than by the needs of the enzyme. Elevated temperatures or pH's outside the range pH 5-9 should only be used in extraordinary circumstances.
- Enzyme treatments should be performed only by persons comfortable interconverting concentration units such as molarity, mM, ppm and (w/v)%, or at least comfortable following the sample calculations given earlier.
- Use only high purity enzymes, usually described in catalogs as crystalline or lyophilized. Use of impure enzyme preparations, usually described as crude, results in adhesion of contaminant proteins that should never have been in the solution in the first place. Crude preparations may even be contaminated with dangerous rogue enzymes like cellulase.
- Increasing treatment temperature may result in a three-to-four-fold increase in rate of reaction, but does so at the cost of considerable increase in logistical difficulty and potential damage to the object.
- Use buffers only for treatments around ph=7. Use acetate buffers rather than phosphate buffers, as the phosphates will precipitate out calcium ions needed by fungal and bacterial enzymes.
- Minimize protein adhesion by using the lowest reasonable concentration of that enzyme, but balance this dictum against potential damage to the object that might result from extended soak times.
- Be sure that adequate concentrations of calcium, sodium and chloride are present in enzyme baths and poultices.
- Remember that denaturation with hot water and/or ethanol is not effective, and that the solvent and/or elevated temperatures may represent a threat to the integrity of the object. Simply rinse instead, preferably with lightly buffered rinse water, though distilled or deionized water works almost as well.
- Washing with ethanol rather than water does not significantly change the amount of protein bound to cellulose, but does change the character of the bound protein.
- Remember that the á-amylase concentration recommendations in this paper are for optimized conditions. Little if any detectable residue is likely to be deposited on the object even at concentrations in the range of 50-250 units activity per ml, as long as in situ denaturation is avoided. If an adhesive is intractable, do not hesitate to increase the α-amylase concentration significantly.
Acknowledgements
The author is deeply indebted to David Laude and Linda Davis, both of whom arranged access to essential equipment, as well as to Karen Pavelka, James Stroud and Olivia Primanis, without whose encouragement and support the work would never have passed outside the walls of the HRHRC.
Citations and Notes
1. Technically, amylases catalyze the hydrolysis of α-D-1,4 glycosidic bonds between α-D-glucopyranose units. An excellent review of the technical details of amylase chemistry can be found in J.F. Robyt, "Enzymes in the Hydrolysis and Synthesis of Starch," in Starch: Chemistry and Technology, 2nd Ed., ed. by R.L. Whistler, J.N. Bemiller and E.F. Paschall, (Orlando: 1984, Academic Press).
2. Pintauro, N.D., Food Processing Enzymes: Recent Developments. (Park Ridge, NJ: Noyes Data Corporation, 1979). W.93-148.
3. Johnson, J.C., Industrial Enzymes: Recent Advances. (Park Ridge, NJ: Noyes Data Corporation, 1977). pp. 148-215.
4. Kulp, K. Carbohydrases, in Food Enzymes in Food Processing, 2nd Ed, ed. by G. Reed. (New York: Academic Press, 1975.) pp. 54-122.
5. Reed, G. Effective temperature and pH, Enzymes in Food Processing 2nd ed. Ed. by G. Reed, (New York: Academic Press, 1975.) pp. 31-42.
6. Gould, B.J., Enzyme data in Handbook of Enzyme Biotechnology, ed. by A. Wiseman. London: Ellis Horwood Limited 1975, pp. 128-162, esp. carbohydrases on pp. 134-145.
7. Wiseman, A., Industrial practice with enzymes, in Handbook of Enzyme Biotechnology, ed. by A. Wiseman. London: Ellis Horwood Limited, 1975, pp. 243-272, esp. carbohydrases on pp. 243 and tables.
8. Daniel, R.M., Bragger, J., Morgan, H.W.; Enzymes from extreme environments, in Biocatalysis, ed. by D.A. Abramowicz. (New York: Van Nostrand Reinhold, 1990). pp. 243-254.
9. Zeikus, J.G., Lowe, S.E., Saha, B.C.; Biocatalysis in anaerobic extremophiles, in Biocatalysis, ed. by D.A. Abramowicz. (New York: Van Nostrand Reinhold 19901. OD. 255-276.
10. Pedersen, S. and Norman, B.E.; Enzymatic modification of food carbohydrates, in Chemical Aspects of Food Enzymes: the proceedings of a symposium organized by The food chemistry group of the industrial division of the Royal Society of Chemistry, ed. by A. T. Andrews. (London: Royal Society of Chemistry, 1987). pp. 156-187.
11. Aunstrup, K., Production, isolation, and economics of extracellular enzymes, in Applied Biochemistry and Bioengineering, Vol.. 2: Enzyme Technology, ed. by Wingard, L.B., Katchalski-Katzir, E. Goldstein, L. (New York: Academic Press, 1979), pp. 27-69.
12. Thoma, J.A., Spradlin, J.E., and Dygert, S; Plant and animal amylases, in The Enzymes, 3rd Ed., Vol. V: Hydrolysis (Sulfate Esters, Carboxyl Esters, Glycosides) and Hydration, ed. by P.D. Boyer. 1971: Academic Press, New York, pp. 115-189. A classic review of the á-amylases of mammals and higher plants.
13. Takagi, T., Toda, H., Isemura, I.; Bacterial and mold amylases, in The Enzymes, 3rd Ed., Vol. V: Hydrolysis (Sulfate Esters, Carboxyl Esters, Glycosides) and Hydration, ed. by P. D. B oyer. 1971: Academic Press, New York pp. 235-271. The classic review of the microbiological á-amylases.
14. G. Bosch, J. Carswell and G. Petherbridge, Islamic Bindings and Bookmaking, The Oriental Institute Museum, The University of Chicago: pp. 35-36.
15. J. Segal and D. Cooper, The Use of Enzymes to Release Adhesives, The Paper Conservator: Journal of The Institute of Paper Conservation, 1977, 2: 47-50.
16. I am grateful to Dr. Greg Wall, Sigma Chemical Company, for the initial suggestion that a test of this sort be developed.
17. A humid oven may be accomplished by maintaining a water level in a pan whose surface area is equal to about half the floor area of the oven. No great harm is inflicted if the pan goes dry occasionally, but the most effective cross-linking will take place in a relatively humid environment. The adhesive may be considered intractable when it no longer releases easily after a brief immersion in water.
18. Cooper, D., King, C., and Segal, J., The use of enzymes in partially non-aqueous media, in The Conservation of Library and Archive Materials and The Graphic Arts: Abstracts and Preprints of the International Conference on the Conservation of Library and Archive Materials arid the Graphic Arts, ed. by G. Petherbridge, (Cambridge: 1980, Institute of Paper Conservation and The Society of Archivists), p. 14-19.
19. Kieboom, A.P.G., Enzymes that do not work in organic solvents: Too polar substrates give too tight enzyme-product complexes, in Biocatalysis, ed. by D.A. Abramowicz. (New York: Van Nostrand Reinhold, 1990). pp. 357-364.
20. Full details of the investigations are reported in a forthcoming monograph or book from researchers led by H. Burgess at the Canadian Conservation Institute, Ottawa The results have been verbally communicated at several venues and are widely accepted. H. Burgess, personal communication.
21. Wakim, J., Robinson, M. and Thoma J.A.; The active site of porcine pancreatic á-amylase: Factors contributing to catalysis, Carbohydrate Research 10, 1969, pp. 487-503, esp. p. 494.
22. Toda H. and Narita, K, Replacement of the essential calcium in Taka-amylase A with other divalent cations, Journal of Biochemistry (Tokyo) 62 1967, pp. 767-768.
23. Prices for manually operated micropipettes range from about $65-$225 ($US, 1992), depending on features, precision, accuracy and service life. The price-conscious occasional user might with reasonable confidence select a micropipette like those described by Cole-Parmer as "Low Cost Pipettes, Adjustable Volume," Cats. No. L-2480-00 (10-50 ìl) and L-24800-10 (50-200 ìl), along with a box of their corresponding disposable tips, Cat. No. L-07953-20 (960 nonsterile pipette tip, 1-200 ìl, prepacked in 10 dispensing racks). Alternatively, users affiliated with a university may wish to borrow a micropipette of the appropriate volume range from a research group in the biological or biochemical sciences.
24. Do not contaminate the bulk liquid by attempting to measure its volume directly. Instead, estimate the volume visually, perhaps by filling a comparable-sized bottle to the same level and measuring the volume of this substitute sample. For the purposes of enzyme preparations, an accuracy of +25% should suffice in any treatment.
25. Dr. Greg Wall Sigma Chemical Co. Technical Support, telephone communication.
26. Anyone extending these principles to proteases should definitely freeze premeasured aliquots of proteases, because refrigerated are subject to autodigestion.
27. Segal and Cooper, 1977, op cit, p. 47
28. personal communication, June 3, 1992.
29. Boel, E., Brady, L, ct al. Calcium binding in á -amylases: An x-ray diffraction study at 2.1~ resolution of two enzymes from aspergillis, Biochemistry 1990, 29, 6244-6249.
30. Thoma, J.A., Spradlin, J.E., Dygert, S; 1971, op cit, p. 133-4.
31. Wakim, J., Robinson, M. and Thoma J.A.; 1969, op cit.
32. Boel, E., Brady, L., ct al, op cit.
33. Lot 50H0607 was labeled by Sigma as containing 1400 units of activity/mg solid and 1900 units/mg protein, where the second figure is merely the result of a Biuret assay of the solid's protein content. Sigma defines a unit of activity as the liberation of "1.0 mg of maltose from starch in 3 minutes at pH 6.9 at 20°C" for most of their á-amylases (Sigma Co. catalog, 1990-91).
34. J. Segal and D. Cooper, 1977, op cit. While Segal and Cooper wed an 0.1%(w/v) solution of á -amylase dissolved in a 10%(V/V) solution of ph=7.2 phosphate buffer, informal polling of conservators indicated that essentially every bench conservator using enzymes was doing so with · solution in straight distilled water. The experiments reported in this article were intended to quantify residues left under treatment conditions actually in use, and were consequently executed with simple aqueous 0.1%(w/v) amylase solutions.
Harold M. EricksonConservation Dept., Harry Ransom Humanities Research Center
University of Texas at Austin
Publication History
Received: Fall 1992
Paper delivered at the Book and Paper specialty group session, AIC 20th Annual Meeting, June 2-7, 1992, Buffalo, NY.
Papers for the specialty group session are selected by committee, based on abstracts and there has been no further peer review. Papers are received by the compiler in the Fall following the meeting and the author is welcome to make revisions, minor or major.